Er heißt Thomas, er ist 38 Jahre alt, und er stellt mir eine Frage, die ich immer häufiger höre: « Mein Vater ist zuckerkrank, meine Mutter hatte Brustkrebs, mein Großvater ist mit 60 Jahren an einem Herzinfarkt gestorben. Ist das in meinen Genen festgeschrieben? Kann ich etwas dagegen tun? » Die Antwort lautet ja. Und nein. Ja, er hat genetische Veranlagungen. Nein, diese Veranlagungen sind keine Verurteilung. Denn zwischen dem Gen und der Krankheit liegt ein ganzer Kontinent, den die Wissenschaft seit zwanzig Jahren erforscht: die Epigenetik. Und diesen Kontinent durchquerst du drei Mal am Tag, bei jedem Essen.
Die Epigenetik ist wohl die wichtigste Entdeckung der Biologie des 21. Jahrhunderts. Sie besagt folgendes: Deine Gene ändern sich nicht, aber sie können durch chemische Modifikationen an oder aus geschaltet werden, die die DNA-Sequenz nicht verändern. Diese Modifikationen sind empfindlich gegenüber der Umwelt: Ernährung, Stress, Giftstoffe, körperliche Bewegung. Und einige sind vererbbar und werden an die nächsten Generationen weitergegeben. Die DNA ist die Partitur. Die Epigenetik ist die Interpretation. Und du bist der Dirigent.
« Epigenetik ist die Untersuchung der Prozesse, durch die der Genotyp, die Gesamtheit der Gene in Wechselwirkung mit der Umwelt, den Phänotyp erzeugt. » Conrad Waddington, 1942
DNA ist nicht nackt
Um die Epigenetik zu verstehen, muss man verstehen, wie DNA im Kern deiner Zellen angeordnet ist. Jede menschliche Zelle enthält 2 Meter DNA (6 Milliarden Basenpaare, jedes mit einem Abstand von 0,34 Nanometer), komprimiert in einem Kern mit einem Durchmesser von 10 Mikrometern. Die Gesamtlänge der DNA eines menschlichen Körpers (50 Billionen Zellen) übersteigt 100 Billionen Meter: das ist 670 Mal die Entfernung Erde-Sonne1. Wie passt man 2 Meter in einen Raum von 10 Mikrometern? Durch Chromatin.
DNA ist niemals nackt. Sie ist um Proteine namens Histone gewickelt. Acht Histone (zwei Kopien von H2A, H2B, H3 und H4) bilden ein Oktamer, um das sich eine Schleife von 146 Nukleotiden DNA wickelt. Das Ganze: Oktamer plus DNA: heißt Nukleosom2. Das ist die Elementarperle der Chromatinkette. Die so gebildete Faser von 11 Nanometern ist das Euchromatin: eine entpackte, zugängliche Struktur, in der Gene transkribiert werden können (gelesen, exprimiert, in Proteine übersetzt). Wenn sich das Histon H1 bindet und die Nukleosomen in einer Solenoidform anordnet, entsteht eine Faser von 30 Nanometern, das Heterochromatin: kompakt, unzugänglich, transkriptional inaktiv3. Die Gene darin sind zum Schweigen gebracht.
Das Heterochromatin ist von zwei Arten. Das konstitutive Heterochromatin ist stabil, irreversibel, in allen Zellen vorhanden (zentromere und telomere Regionen). Das fakultative Heterochromatin ist reversibel: es enthält Gene, die in einigen Zelltypen aktiv und in anderen inaktiv sind4. Dieses fakultative Heterochromatin ist das Spielfeld der Epigenetik. Und hier kommt die Ernährung ins Spiel.
Der Histon-Code: eine biochemische Sprache
Histone sind keine einfachen inerten Spulen. Ihre N-terminalen Enden ragen aus dem Nukleosom heraus und sind das Ziel von post-translationalen Modifikationen, die einen echten biochemischen Code bilden: den Histon-Code5. Diese Modifikationen umfassen die Acetylierung (Addition einer Acetylgruppe durch HAT: Histon-Acetyltransferasen), die Methylierung (Addition einer Methylgruppe durch HMT: Histon-Methyltransferasen), die Phosphorylierung (Addition eines Phosphats durch Kinasen), die Ubiquitinylierung, die SUMOylierung, die Glykosylierung und sogar die Biotinylierung (Addition von Biotin, also von Vitamin B8)6.
Jede Modifikation ist reversibel. HAT acetylieren Lysine. HDAC (Histon-Deacetylasen) deacetylieren sie. HMT methylieren. Demethylasen demethylieren. Professor Laure Weill verwendet in ihrem Kurs zum DU Mikroernährung die Metapher der Writer (diejenigen, die die Markierungen schreiben), der Eraser (diejenigen, die sie löschen) und der Reader (diejenigen, die sie lesen und eine Aktion auslösen)7.
Die Acetylierung der Lysine der Histone H3 und H4 öffnet das Chromatin. Lysine sind bei neutralem pH positiv geladen und interagieren stark mit DNA (negativ geladen). Wenn HAT sie acetylieren, wird die positive Ladung neutralisiert, die Bindung an DNA lockert sich, DNA wird für Transkriptionsfaktoren zugänglich: Gene werden exprimiert8. Die Trimethylierung von H3K9 hingegen rekrutiert das Protein HP1 (Heterochromatin Protein 1), das das Chromatin schließt und die Heterochromatinisierung Schritt für Schritt ausbreitet9. Gleiches Lysin, unterschiedliche Modifikationen, entgegengesetzte Effekte. Das ist der Code.

DNA-Methylierung: wenn Gene zum Schweigen gebracht werden
Die zweite große epigenetische Modifikation ist die Methylierung der DNA selbst. DNA-Methyltransferasen (DNMT) fügen eine Methylgruppe (-CH3) auf Cytosinen hinzu, die sich in CpG-Dinukleotiden befinden (ein C gefolgt von einem G). Diese CpG-Dinukleotide sind oft in « CpG-Inseln » in den Promotorenregionen von Genen gruppiert10. Wenn die CpG-Inseln eines Promoters methyliert sind, ist das Gen zum Schweigen gebracht.
Der Mechanismus ist elegant. Methylierte CpGs rekrutieren Proteine mit MBD (Methyl-CpG Binding Domain) wie MeCP2. MeCP2 rekrutiert HDAC, die benachbarte Histone deacetylieren, und HMT, die H3K9 methylieren. Methyliertes H3K9 rekrutiert HP1. HP1 rekrutiert weitere DNMT. Und die Schleife schließt sich: DNA-Methylierung führt zur Chromatinverdichtung, die zu weiterer Methylierung führt11. Das Schweigen breitet sich aus und wird stabil aufrechterhalten, auch bei der Zellteilung, da DNMT1 methylierte CpGs auf dem elterlichen Strang erkennt und die entsprechenden CpGs auf dem neu synthetisierten Strang methyliert12.
Durch diesen Mechanismus bleiben die Zellen deiner Leber Leberzellen und werden nicht zu Neuronen, obwohl sie genau die gleiche DNA enthalten. « Neuronale » Gene sind methyliert und schweigend in der Leber. « Hepatische » Gene sind methyliert und schweigend im Gehirn. Epigenetik ist das Gedächtnis der Zellidentität.
Die Agouti-Mäuse: der Beweis durch Ernährung
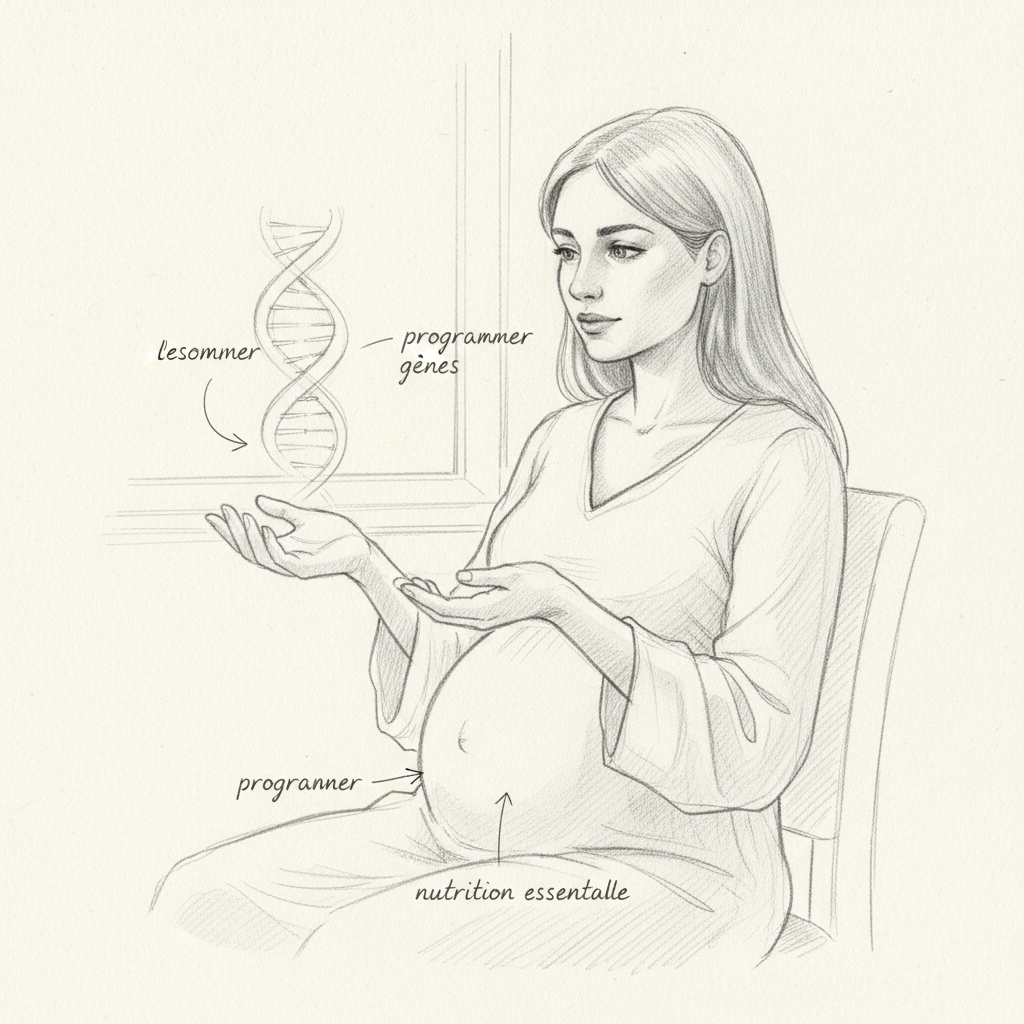
1998 führte Craig Cooney und sein Team aus Arkansas ein Experiment durch, das die Biologie veränderte. Sie nahmen Mäuse, die das Allel Avy (agouti viable yellow) trugen: Ein Transposables Element (IAP), das in das Agouti-Gen eingefügt ist, kontrolliert seine Expression fehlerhaft. Wenn dieser IAP-Promoter aktiv ist, ist die Maus gelb, übergewichtig und hat eine erhöhte Anfälligkeit für Diabetes und Krebs. Alle Mäuse haben genau das gleiche Genom13.
Cooney fütterte zwei Gruppen von trächtigen Mäusen. Gruppe A erhielt Standard-Futter. Gruppe B wurde mit Methylgebern supplementiert: Methionin, Folsäure (Vitamin B9) und Zink: die Vorläufer von S-Adenosylmethionin (SAMe), dem universellen Spender von Methylgruppen im Körper14.
Ergebnis: In Gruppe A (nicht supplementiert) wurden alle Mäuse gelb und übergewichtig geboren. In Gruppe B (mit Methylgebern supplementiert) wurden die meisten Mäuse braun und normalgewichtig geboren. Gleiches Genom. Gleiches Avy-Allel. Aber die mütterliche Ernährung, die die Substrate der Methylierung bereitstellte, hatte die Methylierung der CpG-Inseln des IAP-Promoters verursacht, wodurch die fehlerhafte Expression des Agouti-Gens zum Schweigen gebracht wurde. Die mütterliche Ernährung hatte das Epigenom der Nachkommen neu programmiert.
Das ist nicht Theorie. Das ist experimentelle Biochemie. Und das ist genau das, was die Naturheilkunde seit einem Jahrhundert sagt, wenn sie die Bedeutung der Ernährung während der Schwangerschaft betont. Die Budwig-Creme von Kousmine, reich an B-Vitaminen und Zink, ist keine Marotte. Es ist eine Supplementierung mit Methylgebern.
Die niederländische Hungersnot: wenn Mangel weitergegeben wird
Die Menschheitsgeschichte hat einen tragischen Beweis für die epigenetische transgenerationale Übertragung geliefert. Während des Winters 1944-1945 litten die Niederlande unter einer katastrophalen Hungersnot. Frauen, die während dieser Zeit schwanger waren, bekamen Kinder, die kleiner als normal waren. Aber was Epidemiologen verblüffte, war, dass die Enkel dieser Frauen ebenfalls kleiner als der Durchschnitt waren, obwohl sie nie Hunger erlebt hatten15.
Die Erklärung ist epigenetisch. Der Mangel an Methylgebern (Folsäure, B12, Methionin, Zink) während der Schwangerschaft führte zu einer Abnahme der DNA-Methylierung auf der IGF-2-Gen-Ebene (Insulin-ähnlicher Wachstumsfaktor 2) in der differentiell methylierten Region RDM1. Die Hypomethylierung von RDM1 veränderte die Expression von IGF-2, reduzierte das fetale Wachstum. Und diese Modifikation wurde über das elterliche Prägung an die nächste Generation weitergegeben16. Die Hungersnot der Großmutter hatte eine Markierung in der DNA ihrer Enkel hinterlassen.
Stress, Bienen und Zwillinge
Epigenetik betrifft nicht nur die Ernährung. Stress verändert auch das Epigenom. Experimente mit Kreuzadoptionen bei Ratten (Meaney und Szyf, McGill University) haben dies spektakulär gezeigt. Junge Ratten von gestressten Müttern (wenig Lecken, schwacher Mutterinstinkt), die von ruhigen Müttern adoptiert wurden (häufiges Lecken), werden ruhig. Junge Ratten von ruhigen Müttern, die von gestressten Müttern adoptiert wurden, werden gestresst17. Der Phänotyp « gestresst » ist nicht genetisch. Er ist epigenetisch. Die postnatale Umgebung modifiziert die Methylierungsmarken auf Glukokortikoidrezeptoren im Hippocampus und programmiert die HPA-Achse (Hypothalamus-Hypophysen-Nebennieren-Achse) des Tieres dauerhaft neu.
Bei der Biene ist der Beweis noch eindrucksvoller. Die Larve, die 21 Tage lang mit Gelée Royale gefüttert wird, wird eine fruchtbare Königin, die 3-5 Jahre lebt. Die Larve mit identischem Genom, die 3 Tage lang mit Gelée Royale und dann mit Honig und Pollen gefüttert wird, wird eine unfruchtbare Arbeiterin, die nur wenige Wochen lebt. Gelée Royale hemmt DNMT3 (die de-novo-DNA-Methyltransferase). Von 10.000 analysierten Genen sind 560 zwischen Königinnen und Arbeiterinnen unterschiedlich methyliert18. Gleiche DNA. Unterschiedliche Ernährung. Radikal entgegengesetzte biologische Bestimmung.
Studien über eineiige Zwillinge liefern den letzten Beweis. Mit 3 Jahren haben Zwillinge ein identisches Epigenom. Mit 50 Jahren haben ihre Epigenome sich erheblich unterschieden19. Ernährung, Stress, Giftstoffe, körperliche Aktivität, Infektionen: fünfzig Jahre unterschiedliche Umwelt haben unterschiedliche epigenetische Markierungen auf die gleiche DNA geschrieben. Deshalb kann ein Zwilling Krebs entwickeln und der andere nicht. Es ist nicht die Genetik. Es ist die Epigenetik.

Epigenetik und Krebs: wenn die Sperren brechen
Krebs ist eine epigenetische ebenso wie genetische Krankheit. Professor Weill identifiziert zwei Arten von epigenetischen Modifikationen in Krebszellen20. Zunächst eine globale Hypomethylierung der wiederholten Regionen und Transposable Elements der DNA: Diese Demethylierung verursacht genetische Instabilität (Chromosomenumlagerungen, Translokationen) und die Aktivierung von Onkogenen, die normalerweise stumm sind. Darüber hinaus eine spezifische Hypermethylierung der Promotoren von Tumorsuppressor-Genen, Zellzyklus-Genen, DNA-Reparatur und Apoptose-Genen: Diese Schutzgene sind zum Schweigen gebracht.
Der große Vorteil dieser Entdeckung ist, dass epigenetische Modifikationen reversibel sind. Im Gegensatz zu genetischen Mutationen, die endgültig sind, kann Methylierung entfernt und Histone können wieder acetyliert werden. Das ist die Grundlage der epigenetischen Krebstherapien (DNMT-Inhibitoren, HDAC-Inhibitoren) und auch die Grundlage der ernährungsbasierten Prävention: Ein optimaler Status in Methylgebern könnte vor der globalen Demethylierung schützen, die das Genom destabilisiert.
Die Methylgeber: dein Reprogrammierungs-Kit
S-Adenosylmethionin (SAMe) ist der universelle Spender von Methylgruppen im Körper. Sie wird aus Methionin (essenzielle Aminosäure) durch Methionin-Adenosyltransferase in Gegenwart von ATP hergestellt. Nachdem SAMe seine Methylgruppe gegeben hat, wird sie zu S-Adenosylhomocystein und dann zu Homocystein. Homocystein wird durch zwei Wege zu Methionin recycelt: Der Weg der Methionin-Synthase (die Vitamin B12 als Cofaktor und Methyltetrahydrofolat, abgeleitet von Vitamin B9, als Methyldonor benötigt) und der Weg der Betain-Homocystein-Methyltransferase (die Betain verwendet, das von Cholin stammt)21.
Dieser Methylierungszyklus ist derselbe, den ich im Artikel über die hepatische Entgiftung beschreibe. Er benötigt: Methionin (Fleisch, Fisch, Eier, Paranüsse), Vitamin B9 (Folsäure in Form von Methylfolat: Spinat, Spargel, Leber, Hülsenfrüchte), Vitamin B12 (Methylcobalamin: Fleisch, Fisch, Eier: In pflanzlichen Lebensmitteln nicht enthalten), Zink (Cofaktor der Methionin-Synthase, detailliert im Artikel über Zink), und Cholin/Betain (Eier, Leber, Rüben).
Ein Mangel an einem dieser Nährstoffe beeinträchtigt den Methylierungszyklus, reduziert die Verfügbarkeit von SAMe und verändert möglicherweise die Expression von Hunderten von Genen durch Hypomethylierung. Das ist die biochemische Erklärung des Agouti-Maus-Experiments. Das ist die biochemische Erklärung der niederländischen Hungersnot. Und das ist die biochemische Erklärung der entscheidenden Bedeutung der präkonzeptionellen Supplementierung mit B9 und B12.
Was Thomas verstanden hat
Thomas ist nicht durch die Gene seines Vaters, seiner Mutter und seines Großvaters verurteilt. Seine Gene sind seine Partitur. Aber die Art und Weise, wie diese Partitur gespielt wird, hängt von seiner Ernährung, seinem Lebensstil, seiner Stressmanagement und seinem Status in Methylgebern ab. Seine Bilanz zeigte ein Homocystein von 13 µmol/L (Methylierungsdefizit), niedriges Serum-Zink und eine fast völlige Abwesenheit von grünem Gemüse (Folate) und Fisch (B12, Omega-3) in der Ernährung.
Das Protokoll war einfach und kraftvoll: Behebung des B9-Mangels (Methylfolat 400 µg/Tag), B12 (Methylcobalamin 1000 µg/Tag), Zink-Bisglycinat (25 mg/Tag), und Cholin (über zwei Eier pro Tag und Sonnenblumen-Lecithin). Ernährung reich an grünem Gemüse, Hülsenfrüchten, kleinen fetten Fischen. Stressabbau durch Herz-Kohärenz. Wie ich im Artikel über Zellalterung ausführe, sind Sirtuine, die durch NAD aktiviert werden, selbst große epigenetische Regulatoren (Histon-Deacetylasen). Und vor allem die Einsicht, dass seine täglichen Entscheidungen nicht unbedeutend sind: Sie schreiben jeden Tag die epigenetischen Markierungen, die bestimmen, ob die Gene der Anfälligkeit für Diabetes, Krebs und Herz-Kreislauf-Erkrankungen an- oder ausgeschaltet werden.
Epigenetik sagt nicht, dass Gene nicht zählen. Sie sagt, dass Gene nicht das Schicksal sind. Sie sagt, dass die Umwelt: und an erster Stelle die Ernährung: die Kraft hat, die Genexpression in dieser Generation und in den folgenden zu reprogrammieren. Das ist vielleicht die kraftvollste Botschaft der gesamten modernen Biologie. Und es ist eine Botschaft der Hoffnung.
Du möchtest deinen Status beurteilen? Mache das kostenlose Vitamin-B12-Fragebogen in 2 Minuten.
Um tiefer einzusteigen
- Vitamin B12 (Cobalamin): Methylierung, Neurologie und perniziöse Anämie
- Vitamin B9 (Folate): Methylierung, Schwangerschaft und Homocystein
- Anämie: Tiefe Ursachen verstehen und natürlich handeln
- Krebs und Ernährung: Was Mikroernährung in der Gleichung verändert
Um gesund zu kochen, bietet PranaCook Edelstahlutensilien ohne endokrine Disruptoren an.
Footnotes
-
Weill L. Genetik und Epigenetik. DU Mikroernährung (MAPS). Folie 15: « DNA-Länge in 1 Zelle = 2 Meter. Gesamtlänge = 670 × die Entfernung Sonne-Erde. » ↩
-
Weill L. DU Mikroernährung. Folien 18-19: « Nukleosom: Oktamer (2×H2A, H2B, H3, H4) + 146 Nukleotide DNA. 1. Kompaktionsstufe, 11 nm. » ↩
-
Weill L. DU Mikroernährung. Folien 21-22: « 2. Kompaktionsstufe: H1 → Solenoid 30 nm → Heterochromatin. » ↩
-
Weill L. DU Mikroernährung. Folien 26-27: « Konstitutives Heterochromatin (stabil, irreversibel) vs. fakultatives (reversibel, zelltyp-abhängig). » ↩
-
Weill L. DU Mikroernährung. Folien 30-35: « Der Histon-Code: Kombinationen von Modifikationen kontrollieren die Genexpression. » ↩
-
Weill L. DU Mikroernährung. Folie 34: « Biotinylierung: Addition von Biotin (Vitamin B8). » ↩
-
Weill L. DU Mikroernährung. Folie 38: « Writer (HAT, HMT), Eraser (HDAC, Demethylase), Reader (HP1, Polycomb). » ↩
-
Weill L. DU Mikroernährung. Folie 42: « Acetylierung von Lysinen: Neutralisierung der positiven Ladung → DNA zugänglich → Aktivierung der Transkription. » ↩
-
Weill L. DU Mikroernährung. Folie 43: « Trimethylierung H3K9 → Rekrutierung von HP1 → Ausbreitung der Heterochromatinisierung. » ↩
-
Weill L. DU Mikroernährung. Folien 48-49: « Methylierung der Cytosine auf den CpG-Inseln → Transkriptionsrepression. » ↩
-
Weill L. DU Mikroernährung. Folien 52-53: « MBD → HDAC → HMT → HP1 → DNMT → Ausbreitung und Aufrechterhaltung repressiver epigenetischer Zustände. » ↩
-
Weill L. DU Mikroernährung. Folie 58: « DNMT1 erkennt methylierte Inseln im elterlichen Strang und methyliert die CpG-Inseln im neu synthetisierten Strang. » ↩
-
Weill L. DU Mikroernährung. Folie 73: « Agouti viable yellow Allel: gelbe Maus, übergewichtig, anfällig für Diabetes und Krebs. » ↩
-
Weill L. DU Mikroernährung. Folie 74: « Cooney 1998: Supplementierung mit Methionin, Folsäure und Zink → Methylierung der CpG des IAP-Promoters → braunes Fell, nicht übergewichtig. » ↩
-
Weill L. DU Mikroernährung. Folie 80: « Hungersnot Niederlande 1944-1945: Kinder kleiner als normal, und ihre Kinder auch. Epigenetische Modifikationen werden an nachfolgende Generationen weitergegeben. » ↩
-
Weill L. DU Mikroernährung. Folie 81: « Abnahme der Methylierung auf der IGF-2-Gen-Ebene in der RDM1-Region. » ↩
-
Weill L. DU Mikroernährung. Folien 76-77: « Kreuzadoptionsexperimente: Der gestresste Phänotyp ist epigenetisch, nicht genetisch. » ↩
-
Weill L. DU Mikroernährung. Folie 79: « Gelée Royale hemmt DNMT3. Von 10.000 analysierten Genen sind 560 zwischen Königinnen und Arbeiterinnen unterschiedlich methyliert. » ↩
-
Weill L. DU Mikroernährung. Folie 83: « Zwillinge 3 Jahre: identisches Epigenom. Zwillinge 50 Jahre: divergente Epigenome. » ↩
-
Weill L. DU Mikroernährung. Folie 85: « Krebs: globale Hypomethylierung → genetische Instabilität + spezifische Hypermethylierung → Stummschalten von Tumorsuppressor-Genen. » ↩
-
Weill L. DU Mikroernährung. Folie 74 + Biochemie der Methylierung: « SAMe = universeller Spender. Benötigt Methionin, B9, B12, Zink. » ↩


Laisser un commentaire
Sois le premier à commenter cet article.