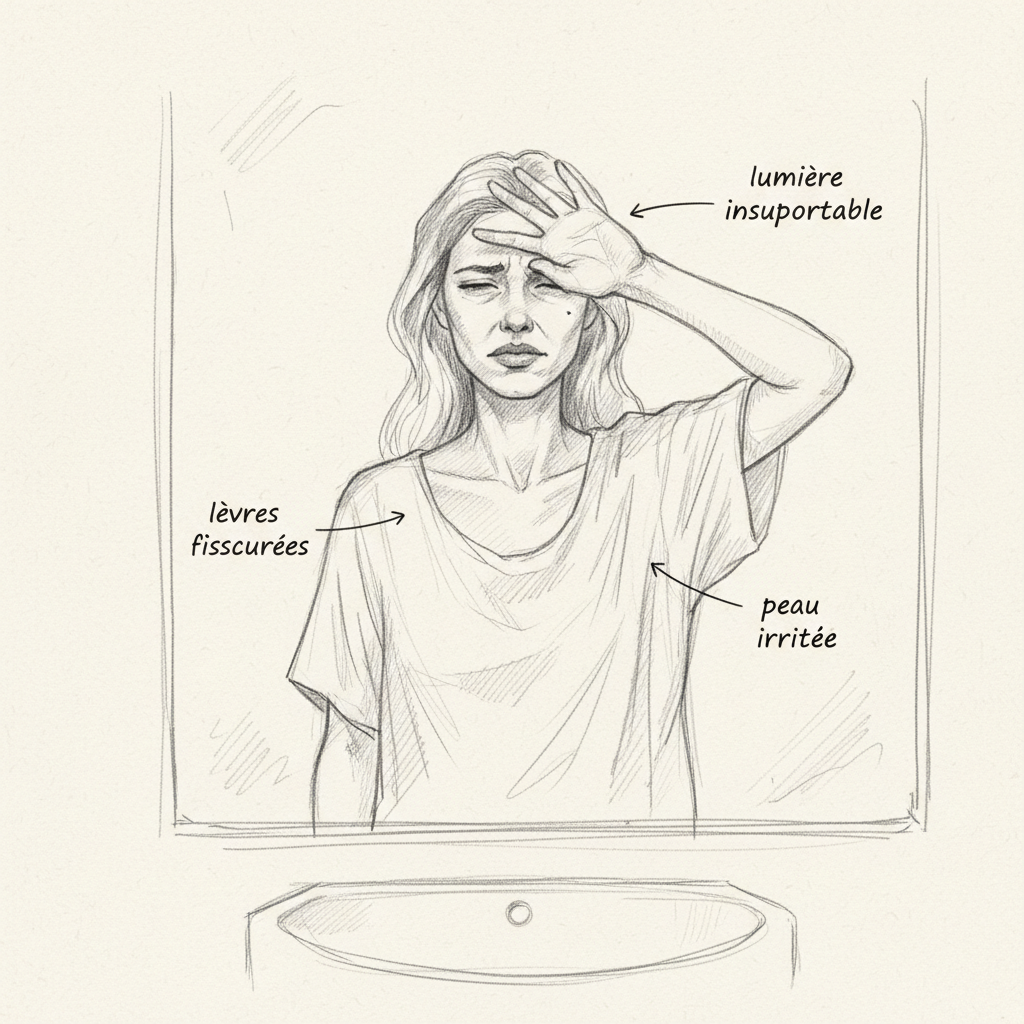
Natália tem quarenta e cinco anos. Ela veio me consultar por uma fadiga crônica que seu médico atribuía ao estresse, uma pele seca que sua dermatologista tratava com creme hidratante, e infecções de ouvido, nariz e garganta repetidas para as quais lhe prescreviam antibióticos a cada dois meses. Investigando sua história clínica, um detalhe me chamou atenção: há aproximadamente um ano, ela tinha dificuldade em dirigir à noite. Os faróis dos carros em sentido contrário a ofuscavam mais, e principalmente, levava um tempo anormalmente longo para recuperar a visão após ser ofuscada. “Meu oftalmologista disse que era normal na minha idade.” Não, não é normal aos quarenta e cinco anos. É um sinal clássico de deficiência de vitamina A, e os médicos antigos sabiam disso.
Examinando seu histórico mais atentamente, constatei que ela tomava um inibidor da bomba de prótons havia três anos para refluxo gástrico, havia se submetido a uma colecistectomia (remoção da vesícula biliar) cinco anos atrás, e comia muito pouca gordura animal por medo do colesterol. Três fatores que convergem para o mesmo resultado: uma má absorção das vitaminas lipossolúveis, incluindo a vitamina A. Seu fígado, privado de bile adequadamente concentrada, não conseguia mais absorver nem armazenar retinol. Sua pele, suas mucosas, seus olhos e seu sistema imunológico pagavam o preço havia meses.
“O terreno é tudo, o micróbio nada.” Antoine Béchamp
A vitamina A é a grande esquecida da micronutrição moderna. Todo mundo fala de vitamina D, magnésio, zinco. Mas a vitamina A, aquela sem a qual você não vê no escuro, aquela que reveste e protege cada mucosa do seu corpo, aquela que permite que seus linfócitos se diferenciem corretamente, é sistematicamente ignorada nos avaliações nutricionais. Ainda assim, a Organização Mundial da Saúde considera a deficiência de vitamina A como a principal causa evitável de cegueira no mundo, e como um fator maior de mortalidade infantil por infecções nos países em desenvolvimento[^1]. Nos países industrializados, a deficiência severa é rara, mas o déficit subclínico é muito mais frequente do que se pensa, especialmente em pessoas que absorvem mal as gorduras.

As causas da deficiência de vitamina A
A vitamina A é uma vitamina lipossolúvel. Essa palavra muda tudo. Lipossolúvel significa que ela precisa de gorduras alimentares para ser absorvida no intestino, de sais biliares para ser emulsificada, de um fígado funcional para ser armazenado, e de proteínas de transporte específicas para circular no sangue. Cada elo dessa corrente pode quebrar, e quando quebra, a deficiência se instala insidiosamente.
A primeira causa, a mais frequente nos países desenvolvidos, é a má absorção de gorduras. Tudo que reduz a secreção biliar ou prejudica a absorção lipídica compromete diretamente a absorção de vitamina A. A insuficiência biliar, seja ligada a uma colecistectomia (como em Natália), litíase biliar, esteatose hepática ou fígado sobrecarregado, reduz drasticamente a capacidade de emulsificação de gorduras no duodeno. Sem bile, as micelas não se formam, e a vitamina A permanece na luz intestinal em vez de ser absorvida pelos enterócitos[^2]. Doença celíaca, doença de Crohn, fibrose cística, pancreatite crônica e síndrome do intestino curto são patologias que prejudicam essa absorção. É exatamente o mesmo mecanismo que afeta a absorção de vitamina D e vitamina E, porque todas as vitaminas lipossolúveis compartilham as mesmas vias de absorção.
A segunda causa é o déficit de ingestão alimentar. Aqui temos duas formas distintas de vitamina A. O retinol pré-formado (ou vitamina A ativa) está presente exclusivamente em produtos animais: fígado, óleo de fígado de bacalhau, manteiga, gema de ovo, queijos gordos. É a forma diretamente utilizável pelo corpo. O beta-caroteno (ou pró-vitamina A) está presente em vegetais coloridos: cenoura, batata-doce, espinafre, manga, damasco. Mas o beta-caroteno não é vitamina A. É um precursor que deve ser convertido em retinol pela enzima BCO1 (beta-caroteno 15,15’-monooxigenase) no enterócito. E essa conversão é muito ineficiente: em média são necessários 12 microgramas de beta-caroteno alimentar para produzir 1 micrograma de retinol[^3]. É uma proporção de 12 para 1. Em algumas pessoas, é ainda pior.
A terceira causa é um fator que a medicina convencional ignora quase completamente: os polimorfismos genéticos da enzima BCO1. Aproximadamente 45% da população europeia carrega uma ou duas variantes do gene BCO1 que reduzem a conversão de beta-caroteno em retinol de 30 a 70%. Essas pessoas são o que a pesquisa chama de “low converters”. Elas podem comer quilos de cenoura sem nunca obter retinol ativo suficiente. O sinal clínico que procuro na consulta: uma pele que fica amarelada nas palmas das mãos e solas dos pés (carotenoderma) apesar do consumo normal de cenoura. Esse amarelecimento, frequentemente confundido com icterícia hepática, na verdade testemunha um acúmulo de beta-caroteno não convertido. É um sinal indireto de polimorfismo BCO1. Em esses pacientes, apenas o retinol pré-formado de origem animal permite cobrir as necessidades.
A quarta causa é o hipotireoidismo. O hormônio tireoidiano T3 é necessário para a expressão do gene BCO1. Quando a tireoide funciona lentamente, a conversão de beta-caroteno em retinol é comprometida. O Dr. Hertoghe enfatiza em The Hormone Handbook que o hipotireoidismo é uma causa subdiagnosticada de deficiência de vitamina A, e que a carotenoderma (pele amarela) é um sinal clássico de hipotireoidismo em sua prática clínica. O círculo é vicioso, porque a vitamina A é ela mesma um cofator do receptor de T3, como veremos mais adiante.
Os sintomas da deficiência
Os olhos são o primeiro órgão a sofrer com um déficit de vitamina A, e faz sentido quando se entende a bioquímica. A vitamina A, em sua forma retinal (aldeído do retinol), é o cromóforo da rodopsina, o pigmento fotossensível dos bastonetes da retina. Os bastonetes são as células responsáveis pela visão em luz baixa, a visão escotópica. Quando um fóton de luz atinge a rodopsina, o 11-cis-retinal se transforma em todo-trans-retinal, disparando o sinal nervoso. Depois o todo-trans-retinal deve ser reciclado em 11-cis-retinal para que o ciclo recomece. Sem ingestão suficiente de vitamina A, esse ciclo desacelera, e a visão noturna se degrada.
A hemeralopia (do grego hemera, dia, e ops, olho), ou cegueira noturna, é o primeiro sinal clínico da deficiência de vitamina A. Era o sinal que Natália apresentava há um ano sem ninguém fazer a conexão. A hemeralopia é reversível se tratada a tempo. Mas se a deficiência continuar, ela evolui para a xeroftalmia: a conjuntiva e a córnea secam, perdem seu brilho. Manchas brancacentas aparecem na conjuntiva, as manchas de Bitot, constituídas de queratina acumulada e bactérias saprófitas. Sem tratamento, a córnea se ulcera (ceratoma lácia), e a cegueira se torna irreversível. A OMS estima que 250 mil a 500 mil crianças ficam cegas a cada ano por causa da deficiência de vitamina A no mundo[^1].
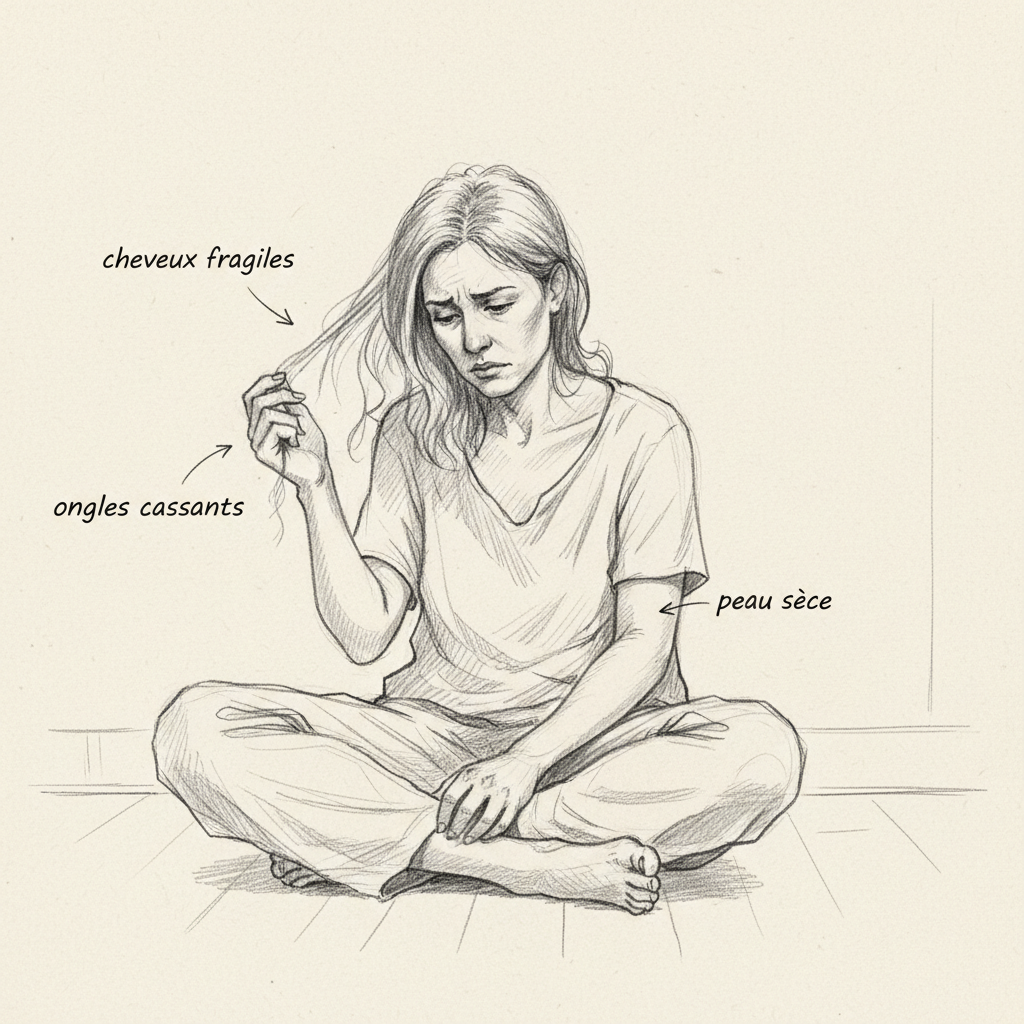
A pele é o segundo espelho da deficiência. A vitamina A, em sua forma ácido retinóico, regula a diferenciação dos queratinócitos, as células que renovam a epiderme. Quando ela falta, a queratinização se torna anárquica: as células da pele se multiplicam sem amadurecer corretamente, e acumulam queratina em excesso. É a hiperqueratose folicular: a pele fica seca, áspera, coberta de pequenas pápulas cornificadas ao redor dos folículos pilosos, especialmente nos braços, coxas e glúteos. Chamam de pele de “carne de galinha permanente”. As mucosas também são afetadas: boca seca, garganta irritada, mucosa vaginal seca, mucosa brônquica frágil. O ácido retinóico é o regulador principal da diferenciação epitelial em todo o corpo. Sem ele, cada superfície mucosa perde sua capacidade de produzir muco protetor e se renovar corretamente.
A imunidade é o terceiro pilar que a deficiência de vitamina A enfraquece. O papel imunológico da vitamina A é duplo e fundamental. Primeiro eixo: a vitamina A é indispensável à diferenciação de linfócitos T ingênuos em linfócitos T reguladores (Treg) no intestino. As células dendríticas da mucosa intestinal produzem ácido retinóico que orienta a maturação dos linfócitos T para um perfil tolerogênico[^4]. Sem vitamina A, os Treg não se formam corretamente, e a imunidade intestinal perde sua capacidade de distinguir o próprio do não-próprio. É um mecanismo que se conecta diretamente à problemática da autoimunidade e da permeabilidade intestinal. Segundo eixo: a vitamina A estimula a produção de IgA secretória, os anticorpos que revestem as mucosas (intestinal, respiratória, urogenital) e constituem a primeira linha de defesa contra agentes patogênicos. Uma deficiência de vitamina A reduz a produção de IgA de 40 a 60% segundo estudos[^5], o que explica por que infecções respiratórias e digestivas são tão frequentes em pessoas deficientes. Natália, com suas otites e bronquites repetidas, ilustrava perfeitamente esse mecanismo.

A vitamina A também desempenha um papel na fertilidade. O ácido retinóico é necessário para a espermatogênese no homem e para o desenvolvimento embrionário normal na mulher. As células de Sertoli nos testículos e o endométrio uterino são tecidos altamente dependentes do ácido retinóico para seu renovação. Em mulheres em idade reprodutiva, um déficit de vitamina A pode contribuir para distúrbios de implantação e abortos precoces, embora esse vínculo ainda necessite mais estudos. A pré-concepção deve incluir uma avaliação do status de vitamina A.
Os micronutrientes essenciais para a vitamina A
A vitamina A nunca trabalha sozinha. Ela se inscreve em uma rede de cofatores cuja deficiência de um compromete o funcionamento dos outros. O mais importante desses cofatores é o zinco. O zinco é indispensável em dois níveis cruciais. Primeiro, é necessário à síntese da RBP (Retinol Binding Protein), a proteína de transporte que veicula o retinol do fígado aos tecidos-alvo. Sem zinco, o retinol permanece sequestrado no fígado e não consegue alcançar as células que dele necessitam[^6]. É uma causa de deficiência funcional de vitamina A mesmo quando as reservas hepáticas são normais. Segundo, o zinco é cofator da retinol desidrogenase, a enzima que converte retinol em retinal na retina. Sem zinco, o ciclo visual da rodopsina é comprometido, e a hemeralopia se instala mesmo com reservas adequadas de vitamina A. O Dr. Curtay, em Nutritherapy, insiste na interdependência zinco-vitamina A: corrigir um sem corrigir o outro é frequentemente insuficiente.
A vitamina D é um parceiro essencial. Os receptores nucleares da vitamina A (RAR e RXR) e da vitamina D (VDR) funcionam em heterodímero: o RXR (receptor do ácido retinóico X) é o parceiro obrigatório do VDR. Em outras palavras, o receptor de vitamina D não consegue funcionar sem um receptor de vitamina A para formar o complexo transcricional ativo. Uma deficiência de vitamina A pode, portanto, comprometer a ação da vitamina D, mesmo que o nível de 25-OH-D3 seja ótimo. É um detalhe bioquímico que muito poucos profissionais conhecem.
As gorduras alimentares são um cofator de absorção indispensável. Como a vitamina A é lipossolúvel, sua ingestão durante uma refeição contendo lipídios (manteiga, azeite de oliva, abacate, oleaginosas) multiplica sua absorção por dois a três. Dietas muito pobres em gorduras, ainda recomendadas por alguns médicos para reduzir o colesterol, são uma causa iatrogênica de má absorção de vitaminas lipossolúveis. A bile é o segundo vetor de absorção: ela emulsifica as gorduras alimentares em micelas, e é dentro dessas micelas que a vitamina A é transportada aos enterócitos. Pessoas que se submeteram a colecistectomia ou sofrem de insuficiência biliar estão, portanto, em risco aumentado.
O ferro participa do metabolismo de vitamina A. A vitamina A melhora a mobilização do ferro das reservas de ferritina e aumenta sua incorporação na hemoglobina. Em retorno, o ferro é necessário para certas enzimas do metabolismo de retinoides. Um estudo de Zimmermann mostrou que a suplementação combinada ferro-vitamina A é mais eficaz que cada uma separadamente para corrigir a anemia em populações deficientes[^7]. É um exemplo concreto de sinergia micronutricional.
As fontes alimentares
O fígado é a fonte mais concentrada em vitamina A pré-formada, de longe. O fígado de bacalhau contém aproximadamente 30 mil microgramas de retinol por 100 gramas, mais de trinta vezes a ingestão diária recomendada. O óleo de fígado de bacalhau contém 18 mil microgramas por 100 gramas. O fígado de boi apporta 9.442 microgramas por 100 gramas. O fígado de aves (frango, peru) contém entre 3.300 e 4.000 microgramas por 100 gramas. Essas concentrações explicam por que um consumo semanal de fígado cobre amplamente as necessidades, e por que povos tradicionais que comiam regularmente miúdos nunca sofriam deficiência de vitamina A. Weston Price, em seus estudos com povos indígenas, observou que as populações mais saudáveis consumiam sistematicamente alimentos ricos em vitaminas lipossolúveis, com o fígado em primeiro lugar.
A manteiga crua (não pasteurizada quando disponível) apporta 750 microgramas por 100 gramas. A gema de ovo contém aproximadamente 380 microgramas por ovo (1.400 microgramas por 100 gramas de gema pura). A gema de ovo tem a vantagem de fornecer simultaneamente vitamina A, vitamina D, colina e fosfolipídios, em uma proporção natural que a indústria nunca conseguiu reproduzir. Os queijos gordos (Emmental, Comté, Roquefort) fornecem entre 250 e 350 microgramas por 100 gramas. Os peixes gordos (enguia, cavala, atum-rabilho) contêm entre 150 e 900 microgramas por 100 gramas dependendo da espécie.
Do lado vegetal, as fontes estão em beta-caroteno, com a proporção de conversão 12:1 a lembrar. A batata-doce alaranjada cozida é a estrela dos carotenoides com aproximadamente 1.000 microgramas de equivalente retinol (ER) por 100 gramas. A cenoura cozida contém 850 microgramas ER por 100 gramas. O espinafre cozido fornece 470 microgramas ER por 100 gramas. A manga contém 54 microgramas ER por 100 gramas. A abóbora-cabotia e a abóbora-manteiga fornecem entre 250 e 400 microgramas ER por 100 gramas. A salsa fresca contém 420 microgramas ER por 100 gramas.
Um ponto essencial: o cozimento suave e a adição de gorduras melhoram consideravelmente a biodisponibilidade do beta-caroteno. O beta-caroteno está aprisionado nas paredes celulares dos vegetais, e o cozimento quebra essas paredes. Uma cenoura cozida com um fio de azeite de oliva fornece três a cinco vezes mais beta-caroteno biodisponível que uma cenoura crua. É um dos casos em que o cozimento suave é preferível ao cru para a assimilação de um nutriente.
Os antagonistas da vitamina A
O álcool é o primeiro antagonista. O etanol e a vitamina A compartilham as mesmas vias de detoxicação hepática, nomeadamente as enzimas álcool desidrogenase (ADH) e aldeído desidrogenase (ALDH). Quando o fígado é mobilizado para metabolizar etanol, não consegue mais converter corretamente retinol em seus metabólitos ativos. O álcool crônico reduz as reservas hepáticas de vitamina A e aumenta simultaneamente a produção de metabólitos tóxicos do retinol. É uma dupla penalidade: menos vitamina A funcional e mais metabólitos prejudiciais. O Dr. Mouton, em Ecologia Digestiva, classifica o álcool crônico como o fator de depleção mais poderoso de vitamina A nos países ocidentais.
Os inibidores da bomba de prótons (IBP) constituem o segundo antagonista. Os IBP, prescritos massivamente para refluxo gástrico, reduzem a acidez gástrica necessária para liberar a vitamina A ligada às proteínas alimentares. Além disso, ao reduzir a acidez duodenal, prejudicam a secreção de colecistoquinina (CCK) que estimula a contração vesicular e a liberação de bile. Menos bile significa menos micelas, e menos micelas significa menos absorção de vitamina A. É o mecanismo encontrado em todas as deficiências de vitaminas lipossolúveis iatrogênicas.
A colestiramina é um sequestrante de ácidos biliares usado para reduzir o colesterol. Ela aprisiona os sais biliares no intestino e impede sua reabsorção. Consequência direta: as gorduras alimentares e as vitaminas lipossolúveis (A, D, E, K) não são mais emulsificadas e atravessam o trato digestivo sem serem absorvidas. Pacientes sob colestiramina de longo prazo desenvolvem deficiências em vitaminas lipossolúveis nos seis a doze meses seguintes ao início do tratamento se nenhuma suplementação for implementada.
O orlistat (Xenical, Alli), um inibidor de lipases pancreáticas prescrito para perda de peso, bloqueia a absorção de trinta por cento das gorduras alimentares. As gorduras não absorvidas levam consigo as vitaminas lipossolúveis. O orlistat reduz a absorção de vitamina A em trinta por cento em média, e os fabricantes recomendam explicitamente suplementação com vitaminas lipossolúveis durante o tratamento. Quantos pacientes sob orlistat nunca receberam essa informação.
O óleo mineral, usado como laxante ou como óleo culinário em certas práticas de emagrecimento, é uma armadilha para vitaminas lipossolúveis. Não sendo absorvido pelo intestino, atravessa o trato digestivo dissolvendo as vitaminas A, D, E e K, que leva nas fezes. É um antagonista mecanicamente terrivelmente eficaz.
A má absorção de gorduras em sentido amplo (insuficiência pancreática exócrina, fibrose cística, ressecção intestinal, doença de Whipple) constitui o último grande antagonista. Qualquer situação que reduza a capacidade do corpo de digerir e absorver lipídios alimentares compromete em cascata a absorção das quatro vitaminas lipossolúveis.
As causas esquecidas da deficiência
A ligação entre vitamina A e tireoide é um assunto que poucos profissionais dominam, e que mereceria por si só um capítulo nos manuais de endocrinologia. O ácido retinóico é um cofator do receptor de T3 (o receptor nuclear TR). O receptor de T3 forma um heterodímero com o RXR, o receptor do ácido retinóico, exatamente como faz o VDR da vitamina D. Sem vitamina A, o sinal de T3 é atenuado no nível genômico, mesmo que os níveis sanguíneos de T3 sejam normais. É uma causa de hipotireoidismo tissular invisível às análises clássicas. Na direção oposta, o hipotireoidismo reduz a atividade de BCO1 e compromete a conversão de beta-caroteno em retinol. O círculo vicioso tireoide-vitamina A é uma armadilha clínica que o Dr. Hertoghe descreve com precisão: corrigir a tireoide melhora o status de vitamina A, e corrigir a vitamina A melhora a resposta tissular a T3.
O microbiota intestinal desempenha um papel pouco conhecido no metabolismo de vitamina A. Estudos recentes mostram que certas cepas bacterianas intestinais (notavelmente Lactobacillus e Bifidobacterium) influenciam a expressão de BCO1 nos enterócitos e modulam a absorção de retinol[^8]. Uma disbiose intestinal severa pode prejudicar o metabolismo de vitamina A independentemente da ingestão alimentar. É mais uma ilustração do papel central do intestino na nutrição, o que a naturopatia afirma há décadas: o primeiro pilar da saúde é a qualidade do seu intestino.
O estresse oxidativo crônico consome vitamina A. O retinol e o beta-caroteno são antioxidantes, e em situação de estresse oxidativo importante (tabagismo, poluição, inflamação crônica, esporte intenso sem recuperação), são consumidos para neutralizar radicais livres, o que reduz a quantidade disponível para suas funções fisiológicas (visão, imunidade, diferenciação celular). É o mesmo mecanismo de “consumo acelerado” observado com zinco em situação de estresse.
O diabetes tipo 2 e a resistência à insulina prejudicam o metabolismo hepático de vitamina A. O fígado esteatósico (fígado gorduroso), extremamente frequente em diabéticos e pessoas com sobrepeso, armazena retinol menos eficientemente e produz menos RBP. As células estreladas hepáticas (células de Ito), que normalmente armazenam oitenta por cento das reservas de vitamina A do corpo, se transformam em miofibroblastos em caso de esteatose e fibrose, perdendo assim sua capacidade de armazenamento. É um mecanismo que Mouton documenta em Ecologia Digestiva: o fígado gorduroso é um fígado que não armazena mais corretamente as vitaminas lipossolúveis.
A interação medicamentosa é uma causa iatrogênica muito frequentemente negligenciada. Além dos IBP, colestiramina e orlistat já mencionados, os retinoides sintéticos (isotretinoína, acitretina) prescritos em dermatologia saturam os receptores RAR e podem paradoxalmente induzir sinais de deficiência funcional de vitamina A natural enquanto geram um excesso de retinoides sintéticos. A metformina, embora não interfira diretamente com vitamina A, pode reduzir a absorção de B12 cujo metabolismo de folatos depende, e os folatos intervêm indiretamente na reciclagem de retinoides. Essas interações em cascata ilustram por que a naturopatia sempre olha o terreno em seu conjunto em vez de um nutriente isolado.
Os suplementos alimentares
A suplementação com vitamina A se faz sob duas formas principais, e a escolha entre as duas depende do contexto clínico.
O palmitato de retinol (ou palmitato de retinila) é a forma pré-formada mais comum em suplemento alimentar. É um éster do retinol que é hidrolisado no intestino para liberar retinol livre, diretamente absorvível. A vantagem do palmitato de retinol é que não depende da enzima BCO1 para ser convertido: é vitamina A ativa. A dosagem comum em adultos é de 800 a 1.500 microgramas (2.600 a 5.000 UI) por dia. A dose máxima tolerada em adultos não grávidas é de 3.000 microgramas (10.000 UI) por dia segundo a EFSA. Acima, o risco de hipervitaminose A (hepatotoxicidade, cefaleia, náuseas, descamação cutânea) aumenta, especialmente em ingestão crônica. Sempre tomar o palmitato de retinol na refeição mais gordurosa do dia para otimizar a absorção.
O beta-caroteno natural é a alternativa vegetal. O beta-caroteno natural, extraído da alga Dunaliella salina ou do óleo de dendê vermelho, contém uma mistura de carotenoides (alfa-caroteno, beta-caroteno, luteína, zeaxantina) que agem sinergeticamente. Distingue-se fundamentalmente do beta-caroteno sintético (todo-trans-beta-caroteno puro) usado em alguns suplementos de baixo custo. O estudo ATBC (Alpha-Tocopherol, Beta-Carotene Cancer Prevention Study) e o estudo CARET mostraram que o beta-caroteno sintético em alta dose (20 a 30 miligramas por dia) aumentava o risco de câncer de pulmão em fumantes, provavelmente por um efeito pró-oxidante paradoxal em alta dose. O beta-caroteno natural em doses fisiológicas (6 a 15 miligramas por dia) nunca demonstrou esse risco. A distinção é capital. A grande vantagem do beta-caroteno é sua segurança: o corpo converte em retinol apenas a quantidade de que precisa, e o excesso é armazenado sem toxicidade (colore a pele em amarelo-alaranjado, só isso). É por essa razão que o beta-caroteno natural é a forma preferida durante a gravidez, onde o risco teratogênico do retinol pré-formado em alta dose é bem documentado. A dose máxima de retinol pré-formado durante a gravidez é de 3.000 microgramas (10.000 UI) por dia, e na prática recomendo não ultrapassar 1.500 microgramas de retinol no primeiro trimestre, privilegiando o beta-caroteno natural para o suplemento.
Para pessoas que têm um polimorfismo BCO1 conhecido ou suspeito (carotenoderma, pele amarela nas palmas apesar de ingestão normal de carotenoides), a suplementação com retinol pré-formado é preferível ao beta-caroteno, já que a via de conversão é deficiente. O bisglicinto de zinco (15 a 30 miligr







Laisser un commentaire
Sois le premier à commenter cet article.